| Weight | 1 lbs |
|---|---|
| Dimensions | 9 × 5 × 2 in |
| host | mouse |
| isotype | IgG1 |
| clonality | monoclonal |
| concentration | 1 mg/mL |
| applications | ICC/IF, WB |
| reactivity | p53 |
| available sizes | 100 µg |



mouse anti-p53 monoclonal antibody (240) 5777
$503.00
Antibody summary
- Mouse monoclonal to p53
- Suitable for: WB,ICC/IF,IHC-P,IHC-Fr,FACS,IP,ELISA
- Isotype: IgG1
- 100 µg
mouse anti-p53 monoclonal antibody (240) 5777
| antibody |
|---|
| Tested applications WB,IHC,IHC,ICC/IF |
| Recommended dilutions ELISA, FACS, IHC (frozen and paraffin), Immunoprecipitation, Immunoblotting. IHC: use at 2-4ug/ml. Extensive washing may be necessary. Protein digestion and/or microwave antigen retrieval prior to staining is not required. #3340 on paraffin-embedded HBL435 xenograft. Immunoprecipita |
| Immunogen Gel-purified p53 containing aa 14-389 of human p53. |
| Size and concentration 100µg and lot specific |
| Form liquid |
| Storage Instructions This antibody is stable for at least one (1) year at -20°C. Avoid multiple freeze-thaw cycles. |
| Storage buffer PBS, pH 7.4 |
| Purity protein affinity purification |
| Clonality monoclonal |
| Isotype IgG1 |
| Compatible secondaries goat anti-mouse IgG, H&L chain specific, peroxidase conjugated polyclonal antibody 5486 goat anti-mouse IgG, H&L chain specific, biotin conjugated, Conjugate polyclonal antibody 2685 goat anti-mouse IgG, H&L chain specific, FITC conjugated polyclonal antibody 7854 goat anti-mouse IgG, H&L chain specific, peroxidase conjugated polyclonal antibody, crossabsorbed 1706 goat anti-mouse IgG, H&L chain specific, biotin conjugated polyclonal antibody, crossabsorbed 1716 goat anti-mouse IgG, H&L chain specific, FITC conjugated polyclonal antibody, crossabsorbed 1721 |
| Isotype control Mouse monocolonal IgG1 - Isotype Control |
| target relevance |
|---|
| Homo sapiens TP53 Cellular tumor antigen p53 |
| Protein names Cellular tumor antigen p53 |
| Alternative names Antigen NY-CO-13, Phosphoprotein p53, Tumor suppressor p53 |
| Gene names TP53 |
| Protein family Belongs to the p53 family |
| Function Multifunctional transcription factor that induces cell cycle arrest, DNA repair or apoptosis upon binding to its target DNA sequence (PubMed:11025664, PubMed:12524540, PubMed:12810724, PubMed:15186775, PubMed:15340061, PubMed:17317671, PubMed:17349958, PubMed:19556538, PubMed:20673990, PubMed:20959462, PubMed:22726440, PubMed:24051492, PubMed:24652652, PubMed:35618207, PubMed:36634798, PubMed:38653238, PubMed:9840937). Acts as a tumor suppressor in many tumor types; induces growth arrest or apoptosis depending on the physiological circumstances and cell type (PubMed:11025664, PubMed:12524540, PubMed:12810724, PubMed:15186775, PubMed:15340061, PubMed:17189187, PubMed:17317671, PubMed:17349958, PubMed:19556538, PubMed:20673990, PubMed:20959462, PubMed:22726440, PubMed:24051492, PubMed:24652652, PubMed:38653238, PubMed:9840937). Negatively regulates cell division by controlling expression of a set of genes required for this process (PubMed:11025664, PubMed:12524540, PubMed:12810724, PubMed:15186775, PubMed:15340061, PubMed:17317671, PubMed:17349958, PubMed:19556538, PubMed:20673990, PubMed:20959462, PubMed:22726440, PubMed:24051492, PubMed:24652652, PubMed:9840937). One of the activated genes is an inhibitor of cyclin-dependent kinases. Apoptosis induction seems to be mediated either by stimulation of BAX and FAS antigen expression, or by repression of Bcl-2 expression (PubMed:12524540, PubMed:17189187). Its pro-apoptotic activity is activated via its interaction with PPP1R13B/ASPP1 or TP53BP2/ASPP2 (PubMed:12524540). However, this activity is inhibited when the interaction with PPP1R13B/ASPP1 or TP53BP2/ASPP2 is displaced by PPP1R13L/iASPP (PubMed:12524540). In cooperation with mitochondrial PPIF is involved in activating oxidative stress-induced necrosis; the function is largely independent of transcription. Induces the transcription of long intergenic non-coding RNA p21 (lincRNA-p21) and lincRNA-Mkln1. LincRNA-p21 participates in TP53-dependent transcriptional repression leading to apoptosis and seems to have an effect on cell-cycle regulation. Implicated in Notch signaling cross-over. Prevents CDK7 kinase activity when associated to CAK complex in response to DNA damage, thus stopping cell cycle progression. Isoform 2 enhances the transactivation activity of isoform 1 from some but not all TP53-inducible promoters. Isoform 4 suppresses transactivation activity and impairs growth suppression mediated by isoform 1. Isoform 7 inhibits isoform 1-mediated apoptosis. Regulates the circadian clock by repressing CLOCK-BMAL1-mediated transcriptional activation of PER2 (PubMed:24051492) |
| Subcellular location Cytoplasm |
| Structure (Microbial infection) Interacts with Kaposi's sarcoma-associated herpesvirus/HHV-8 protein ORF45; this interaction results in the cytoplasmic localization of TP53 thereby decreasing its transcriptional activity |
| Post-translational modification Acetylation of Lys-382 by CREBBP enhances transcriptional activity (PubMed:10656795, PubMed:15448695, PubMed:20228809, PubMed:23431171). Acetylation of Lys-382 by EP300 (PubMed:10656795, PubMed:15448695, PubMed:20228809, PubMed:23431171). Deacetylation of Lys-382 by SIRT1 impairs its ability to induce proapoptotic program and modulate cell senescence (PubMed:10656795, PubMed:15448695, PubMed:20228809, PubMed:23431171). Deacetylation by SIRT2 impairs its ability to induce transcription activation in a AKT-dependent manner (PubMed:10656795, PubMed:15448695, PubMed:20228809, PubMed:23431171, PubMed:29681526). Acetylation at Lys-381 increases stability (PubMed:29474172). Deacetylation at Lys-381 by SIRT6 decreases its stability, thereby regulating cell senescence (PubMed:29474172). Acetylated at Lys-120 by KAT5, KAT6A and KAT8; regulating its ability to induce proapoptotic program (PubMed:17189187, PubMed:19854137, PubMed:23431171) Lactylation by AARS1 prevents ability to undergo liquid-liquid phase separation (LLPS), thereby inhibiting transcription factor activity Phosphorylation on Ser residues mediates transcriptional activation. Phosphorylated by HIPK1 (By similarity). Phosphorylation at Ser-9 by HIPK4 increases repression activity on BIRC5 promoter. Phosphorylated on Thr-18 by VRK1, which may prevent the interaction with MDM2 (PubMed:10951572, PubMed:31527692). Phosphorylated on Ser-20 by CHEK2 in response to DNA damage, which prevents ubiquitination by MDM2. Phosphorylated on Ser-20 by PLK3 in response to reactive oxygen species (ROS), promoting p53/TP53-mediated apoptosis. Phosphorylated on Thr-55 by TAF1, which promotes MDM2-mediated degradation. Phosphorylated on Ser-33 by CDK7 in a CAK complex in response to DNA damage. Phosphorylated on Ser-46 by HIPK2 upon UV irradiation. Phosphorylation on Ser-46 is required for acetylation by CREBBP. Phosphorylated on Ser-392 following UV but not gamma irradiation. Phosphorylated by NUAK1 at Ser-15 and Ser-392; was initially thought to be mediated by STK11/LKB1 but it was later shown that it is indirect and that STK11/LKB1-dependent phosphorylation is probably mediated by downstream NUAK1 (PubMed:21317932). It is unclear whether AMP directly mediates phosphorylation at Ser-15. Phosphorylated on Thr-18 by isoform 1 and isoform 2 of VRK2. Phosphorylation on Thr-18 by isoform 2 of VRK2 results in a reduction in ubiquitination by MDM2 and an increase in acetylation by EP300. Stabilized by CDK5-mediated phosphorylation in response to genotoxic and oxidative stresses at Ser-15, Ser-33 and Ser-46, leading to accumulation of p53/TP53, particularly in the nucleus, thus inducing the transactivation of p53/TP53 target genes. Phosphorylated by DYRK2 at Ser-46 in response to genotoxic stress. Phosphorylated at Ser-315 and Ser-392 by CDK2 in response to DNA-damage. Phosphorylation at Ser-15 is required for interaction with DDX3X and gamma-tubulin (PubMed:28842590). Phosphorylation at Ser-392 regulates its ability to undergo liquid-liquid phase separation by increasing fluidity of TP53/p53 condensates (PubMed:35618207) Dephosphorylated by PP2A-PPP2R5C holoenzyme at Thr-55. SV40 small T antigen inhibits the dephosphorylation by the AC form of PP2A May be O-glycosylated in the C-terminal basic region. Studied in EB-1 cell line Ubiquitinated by MDM2 and SYVN1, which leads to proteasomal degradation (PubMed:10722742, PubMed:12810724, PubMed:15340061, PubMed:17170702, PubMed:19880522, PubMed:29681526). Ubiquitinated by RFWD3, which works in cooperation with MDM2 and may catalyze the formation of short polyubiquitin chains on p53/TP53 that are not targeted to the proteasome (PubMed:10722742, PubMed:12810724, PubMed:20173098). Ubiquitinated by MKRN1 at Lys-291 and Lys-292, which leads to proteasomal degradation (PubMed:19536131). Deubiquitinated by USP10, leading to its stabilization (PubMed:20096447). Ubiquitinated by TRIM24, RFFL, RNF34 and RNF125, which leads to proteasomal degradation (PubMed:19556538). Ubiquitination by TOPORS induces degradation (PubMed:19473992). Deubiquitination by USP7, leading to stabilization (PubMed:15053880). Isoform 4 is monoubiquitinated in an MDM2-independent manner (PubMed:15340061). Ubiquitinated by COP1, which leads to proteasomal degradation (PubMed:19837670). Ubiquitination and subsequent proteasomal degradation is negatively regulated by CCAR2 (PubMed:25732823). Polyubiquitinated by C10orf90/FATS, polyubiquitination is 'Lys-48'-linkage independent and non-proteolytic, leading to TP53 stabilization (By similarity). Polyubiquitinated by MUL1 at Lys-24 which leads to proteasomal degradation (PubMed:21597459). Deubiquitinated by USP3, leading to stabilization (PubMed:28807825). Ubiquitinated by MSL2, promoting its cytoplasmic localization (PubMed:19033443). Also ubiquitinated by the SCF(FBXO22)-KDMA4A complex; leading to proteasomal degradation (PubMed:26868148) Monomethylated at Lys-372 by SETD7, leading to stabilization and increased transcriptional activation (PubMed:15525938, PubMed:16415881). Monomethylated at Lys-370 by SMYD2, leading to decreased DNA-binding activity and subsequent transcriptional regulation activity (PubMed:17108971). Lys-372 monomethylation prevents interaction with SMYD2 and subsequent monomethylation at Lys-370 (PubMed:17108971). Dimethylated at Lys-373 by EHMT1 and EHMT2 (PubMed:20118233). Monomethylated at Lys-382 by KMT5A, promoting interaction with L3MBTL1 and leading to repress transcriptional activity (PubMed:17707234). Dimethylation at Lys-370 and Lys-382 diminishes p53 ubiquitination, through stabilizing association with the methyl reader PHF20 (PubMed:22864287). Demethylation of dimethylated Lys-370 by KDM1A prevents interaction with TP53BP1 and represses TP53-mediated transcriptional activation (PubMed:17805299). Monomethylated at Arg-333 and dimethylated at Arg-335 and Arg-337 by PRMT5; methylation is increased after DNA damage and might possibly affect TP53 target gene specificity (PubMed:19011621) Sumoylated with SUMO1. Sumoylated at Lys-386 by UBC9 |
| Involvement in disease Esophageal cancer A malignancy of the esophagus. The most common types are esophageal squamous cell carcinoma and adenocarcinoma. Cancer of the esophagus remains a devastating disease because it is usually not detected until it has progressed to an advanced incurable stage. Li-Fraumeni syndrome An autosomal dominant familial cancer syndrome that in its classic form is defined by the existence of a proband affected by a sarcoma before 45 years with a first degree relative affected by any tumor before 45 years and another first degree relative with any tumor before 45 years or a sarcoma at any age. Other clinical definitions for LFS have been proposed and called Li-Fraumeni like syndrome (LFL). In these families affected relatives develop a diverse set of malignancies at unusually early ages. Four types of cancers account for 80% of tumors occurring in TP53 germline mutation carriers: breast cancers, soft tissue and bone sarcomas, brain tumors (astrocytomas) and adrenocortical carcinomas. Less frequent tumors include choroid plexus carcinoma or papilloma before the age of 15, rhabdomyosarcoma before the age of 5, leukemia, Wilms tumor, malignant phyllodes tumor, colorectal and gastric cancers. Squamous cell carcinoma of the head and neck A non-melanoma skin cancer affecting the head and neck. The hallmark of cutaneous SCC is malignant transformation of normal epidermal keratinocytes. Lung cancer A common malignancy affecting tissues of the lung. The most common form of lung cancer is non-small cell lung cancer (NSCLC) that can be divided into 3 major histologic subtypes: squamous cell carcinoma, adenocarcinoma, and large cell lung cancer. NSCLC is often diagnosed at an advanced stage and has a poor prognosis. Papilloma of choroid plexus A benign tumor of neuroectodermal origin that generally occurs in childhood, but has also been reported in adults. Although generally found within the ventricular system, choroid plexus papillomas can arise ectopically in the brain parenchyma or disseminate throughout the neuraxis. Patients present with signs and symptoms of increased intracranial pressure including headache, hydrocephalus, papilledema, nausea, vomiting, cranial nerve deficits, gait impairment, and seizures. Adrenocortical carcinoma A malignant neoplasm of the adrenal cortex and a rare childhood tumor. It occurs with increased frequency in patients with Beckwith-Wiedemann syndrome and Li-Fraumeni syndrome. Basal cell carcinoma 7 A common malignant skin neoplasm that typically appears on hair-bearing skin, most commonly on sun-exposed areas. It is slow growing and rarely metastasizes, but has potentialities for local invasion and destruction. It usually develops as a flat, firm, pale area that is small, raised, pink or red, translucent, shiny, and waxy, and the area may bleed following minor injury. Tumor size can vary from a few millimeters to several centimeters in diameter. Bone marrow failure syndrome 5 A form of bone marrow failure syndrome, a heterogeneous group of life-threatening disorders characterized by hematopoietic defects in association with a range of variable extra-hematopoietic manifestations. BMFS5 is an autosomal dominant form characterized by infantile onset of severe red cell anemia requiring transfusion. Additional features include hypogammaglobulinemia, poor growth with microcephaly, developmental delay, and seizures. |
| Keywords 3D-structure, Acetylation, Activator, Alternative promoter usage, Alternative splicing, Apoptosis, Biological rhythms, Cell cycle, Cytoplasm, Cytoskeleton, Direct protein sequencing, Disease variant, DNA-binding, Endoplasmic reticulum, Glycoprotein, Host-virus interaction, Isopeptide bond, Li-Fraumeni syndrome, Metal-binding, Methylation, Mitochondrion, Necrosis, Nucleus, Phosphoprotein, Proteomics identification, Reference proteome, Repressor, Transcription, Transcription regulation, Tumor suppressor, Ubl conjugation, Zinc |
| Sequence MEEPQSDPSVEPPLSQETFSDLWKLLPENNVLSPLPSQAMDDLMLSPDDIEQWFTEDPGP DEAPRMPEAAPPVAPAPAAPTPAAPAPAPSWPLSSSVPSQKTYQGSYGFRLGFLHSGTAK SVTCTYSPALNKMFCQLAKTCPVQLWVDSTPPPGTRVRAMAIYKQSQHMTEVVRRCPHHE RCSDSDGLAPPQHLIRVEGNLRVEYLDDRNTFRHSVVVPYEPPEVGSDCTTIHYNYMCNS SCMGGMNRRPILTIITLEDSSGNLLGRNSFEVRVCACPGRDRRTEEENLRKKGEPHHELP PGSTKRALPNNTSSSPQPKKKPLDGEYFTLQIRGRERFEMFRELNEALELKDAQAGKEPG GSRAHSSHLKSKKGQSTSRHKKLMFKTEGPDSD |
| UniProt accession: P04637 |
Data
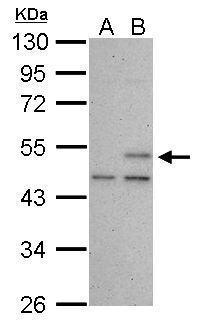 |
| Sample (30 µg of whole cell lysate) A: HCT116 cells with mock treatment for 24 hr B: HCT116 cells with 30 uM cisplatin treatment for 24 hr 10% SDS PAGE 5777 diluted at 1:1000 The HRP-conjugated anti-mouse IgG antibody was used to detect the primary antibody. |
 |
| Immunohistochemical analysis of paraffin-embedded HBL435 xenograft, using p53(5777) antibody at 1:200 dilution. |
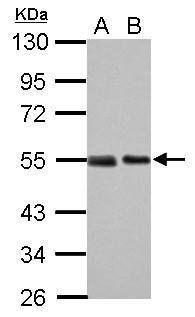 |
| Sample (30 µg of whole cell lysate) A: 293T B: A431 10% SDS PAGE 5777 diluted at 1:1000 The HRP-conjugated anti-mouse IgG antibody was used to detect the primary antibody. |
FAQ & Publications
Frequently Asked Questions
What applications has the mouse anti-p53 monoclonal antibody (240) 5777 been tested and recommended for?
This antibody has been tested and is suitable for Western blotting (WB), immunocytochemistry/immunofluorescence (ICC/IF), immunohistochemistry on paraffin-embedded and frozen tissues (IHC-P, IHC-Fr), flow cytometry (FACS), immunoprecipitation (IP), and ELISA.
How should the mouse anti-p53 monoclonal antibody (240) 5777 be stored to maintain its stability?
The antibody should be stored at -20°C and is stable for at least one year under these conditions. It is recommended to avoid multiple freeze-thaw cycles to preserve antibody integrity.
Publications
| pmid | title | authors | citation |
|---|---|---|---|
| 36636131 | 3D printing biocompatible materials with Multi Jet Fusion for bioreactor applications. | Balasankar Meera Priyadarshini, Wai Kay Kok, Vishwesh Dikshit, Shilun Feng, King Ho Holden Li, Yi Zhang | Int J Bioprint 9:623 |
| 36582449 | Genetically engineered probiotics as catalytic glucose depriver for tumor starvation therapy. | Penghao Ji, Bolin An, Zhongming Jie, Liping Wang, Shuwen Qiu, Changhao Ge, Qihui Wu, Jianlin Shi, Minfeng Huo | Mater Today Bio 18:100515 |
| 36578521 | METTL14 Regulates Intestine Cellular Senescence through m(6)A Modification of Lamin B Receptor. | Zizhen Zhang, Meng Xue, Jingyu Chen, Zhuo Wang, Fangyu Ju, Jiaojiao Ni, Jiawei Sun, Haoyue Wu, Huimei Zheng, Ziwei Lou, Yawen Zhang, Xiaohang Yang, Shujie Chen, Yongmei Xi, Liangjing Wang | Oxid Med Cell Longev 2022:9096436 |
| 36430911 | Transcriptome Analysis of Goat Mammary Gland Tissue Reveals the Adaptive Strategies and Molecular Mechanisms of Lactation and Involution. | Rong Xuan, Jianmin Wang, Xiaodong Zhao, Qing Li, Yanyan Wang, Shanfeng Du, Qingling Duan, Yanfei Guo, Zhibin Ji, Tianle Chao | Int J Mol Sci 23:N/A |
| 36376321 | Replication collisions induced by de-repressed S-phase transcription are connected with malignant transformation of adult stem cells. | Ting Zhang, Carsten Künne, Dong Ding, Stefan Günther, Xinyue Guo, Yonggang Zhou, Xuejun Yuan, Thomas Braun | Nat Commun 13:6907 |
Published literature highly relevant to the biological target of this product and referencing this antibody or clone are retrieved from the PubMed database provided by the United States National Library of Medicine at the National Institutes of Health.
Protocols
| relevant to this product |
|---|
| Western blot IHC ICC |
Documents
| Batch Number | QC File | SDS |
|---|---|---|
| To view batch-specific Safety Datasheets and Quality Certificates associated with your account, please Log In. | ||
Only logged in customers who have purchased this product may leave a review.
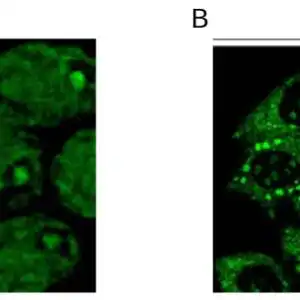



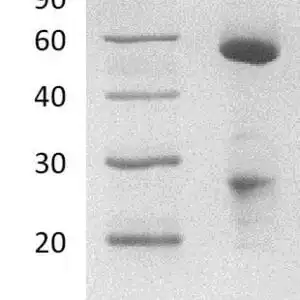

Reviews
There are no reviews yet.